
铝合金霉菌腐蚀研究进展
2022-07-14 16:59:31
作者: 张雨轩, 陈翠颖, 刘宏伟, 李伟华 来源:中国腐蚀与防护学报
分享至:


摘要
结合近年来铝合金霉菌腐蚀机制与防护领域研究成果,介绍了代表性霉菌的种类及影响霉菌活性的主要因素,重点总结和讨论了铝合金霉菌腐蚀机制,主要包括酸蚀机制、浓差电池机制、以及其他可能存在直接电子传递机制和霉菌铝合金直接界面作用机制。霉菌通过新陈代谢可以产生大量的有机酸,能够显著地降低介质和生物膜内的pH,从而导致酸蚀引发局部腐蚀。霉菌在铝合金表面形成的生物膜是诱发氧浓差电池产生的原因之一,铝合金霉菌腐蚀过程中潜在的直接电子传递及铝合金和霉菌的直接界面作用也是导致铝合金局部腐蚀的重要原因之一。最后介绍了目前常用的铝合金霉菌腐蚀控制方法,展望了未来铝合金霉菌腐蚀的研究重点,为铝合金霉菌腐蚀研究提供参考。
关键词: 铝合金; 霉菌; 酸蚀; 微生物腐蚀
铝合金材料具有强度高、密度小、力学性能好、易加工等优点,广泛应用于各种工程材料以及飞机、舰船等高端设备,目前其使用量仅次于钢铁[1,2]。对于民航飞机来说,铝合金的用量一般占到80%左右,在现代舰载机中,铝合金的用量也在60%左右[3]。铝合金在自然条件下会在其表面形成一层厚约10~100 nm的Al2O3膜,具有良好的附着力和自愈合性,能够在pH 4~9的溶液中稳定存在,起到钝化膜作用,对基体铝合金材料具有良好的保护作用[4]。铝合金在大气环境中形成氧化物膜层较薄,疏松多孔且不连续,保护性较差。在沿海高温、高盐、高湿海洋环境中,铝合金装备腐蚀失效问题频发,现场检测腐蚀形态以局部腐蚀为主,其失效机制包括:小孔腐蚀、应力腐蚀开裂、缝隙腐蚀、电偶腐蚀以及微生物腐蚀 (MIC) 等[4,5]。
近年来,研究发现微生物腐蚀是造成结构材料和构件腐蚀失效的一个关键原因之一,海洋环境中微生物腐蚀造成的经济损失占总经济损失的一半以上,因此国内外越来越多研究人员开始关注微生物腐蚀,微生物腐蚀研究的投入也逐渐增大[6-8]。微生物腐蚀是指微生物通过自身生命活动及其代谢产物直接和间接地加速或者抑制金属材料腐蚀过程的现象[9]。微生物腐蚀是一个复杂的电化学过程,微生物腐蚀的发生、发展与微生物的滋生、其在材料表面附着以及微生物新陈代谢产物等直接相关,甚至在有机碳源供应不足的情况下,可以直接从基体金属材料获得电子,为其生长提供能量,通过生物氧化显著地加速金属的腐蚀[10-12]。近年来研究表明,微生物腐蚀也是造成铝合金结构构件服役失效的主要原因之一,其中霉菌是主要的腐蚀性微生物。美国空军的C-130、KC135、B-52机翼内部曾受过芽枝霉和铜绿色极毛杆菌的严重腐蚀,20世纪70年代,我国某飞机系统附件霉变受损,而诱发了飞行事故[13,14]。Christopher等[15]研究发现霉菌是飞机燃料的主要污染微生物,也是造成飞机油箱腐蚀失效的原因之一。Hagenauer等[16]对于飞机腐蚀破坏区域取样分析也表明霉菌是造成7075铝合金快速腐蚀的主要真菌。
霉菌腐蚀是威胁着铝合金结构构件及设备安全服役的关键因素之一,近年来,国内外研究人员围绕着铝合金的霉菌腐蚀机制和防护研究做了大量的工作。本文结合最新的研究成果,系统地总结了造成铝合金服役失效的关键菌群种类、铝合金霉菌腐蚀机制、控制铝合金霉菌腐蚀的措施和方法,从而为后续铝合金霉菌研究和控制提供参考。
1 霉菌的种类及特性
1.1 霉菌种类
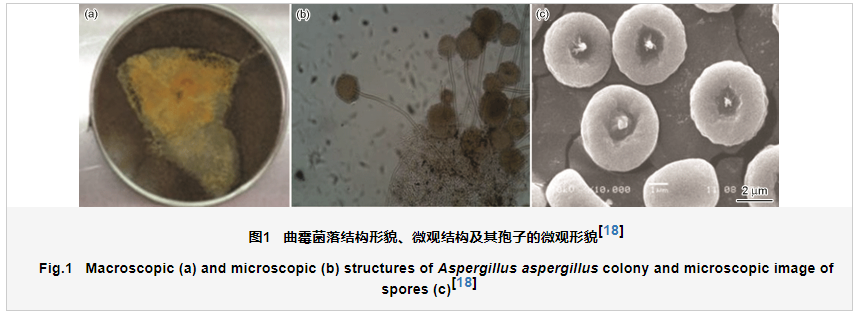
霉菌是丝状真菌的统称,是真核生物,霉菌包含分类学上许多不同纲或类的真菌,分别属于藻状菌纲、子囊菌纲、担子菌纲和半知菌纲等。霉菌代谢活性强,生命力旺盛,广泛存在于大气、土壤、水等各种自然环境中。霉菌通常都是以菌丝进行生长,以孢子进行繁殖。外界环境条件在霉菌适宜生长的温度 (20~40 ℃) 和湿度内 (湿度超过70%),同时如果有充足的碳源、氮源和无机盐等营养物质时,霉菌的孢子可以在各种材料表面快速萌芽和生长。在霉菌的生长过程中,如果缺乏营养物,霉菌在用完孢子内的养料后就会停止生长。研究发现,霉菌的种类大约有10万种,其中影响材料服役过程的霉菌预计约有数万种[17]。代表性霉菌之一的曲霉菌落结构以及其微观结构和孢子如图1,从图中可以看出,霉菌呈丝状,孢子直径在5 μm左右[18]。研究表明,对于铝合金腐蚀性较强的代表性霉菌菌群主要有:曲霉属、青霉属、链格孢属、短梗霉属、木霉属、枝孢属、毛壳菌属等,代表性霉菌主要包括:黑曲霉、米根霉、华根霉、鲁氏毛霉、总状毛霉、米曲霉、黄曲霉、栖土霉菌、产黄青霉、桔青霉等 [19]。霉菌种类的差异会对霉菌的代谢活性以及在材料表面的吸附过程产生影响,从而会直接影响其在材料表面生物膜的形成和结构,对于铝合金腐蚀行为和腐蚀机制也会产生显著的影响。但是霉菌的种类众多,霉菌菌群结构差异对铝合金腐蚀行为和机制的研究还有待深入。
1.2 霉菌的特性
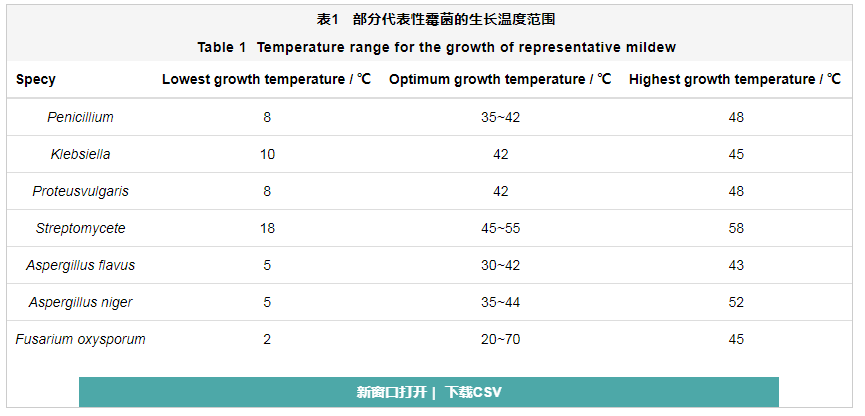
霉菌的主要营养物质包括碳素化合物、氮素化合物、无机盐等。碳源是构成细胞的关键组分,占了霉菌细胞干重的一半以上,是霉菌维持新陈代谢的主要能源之一;氮素营养用于合成氨基酸、核酸和各种维生素;Na、K、Mg、Fe等的无机盐主要用于调节细胞膜的渗透压和维持细胞内酶的活性。温度对霉菌的活性会产生显著的影响,部分代表性霉菌的生长温度范围如表1所示。可以看出,霉菌的最适生长温度在30 ℃左右。南海湿热的环境为霉菌的快速生长创造了良好的自然条件,也间接地促进了铝合金构件的霉菌腐蚀。
霉菌细胞内含有大量的水,含量在58.79%~88.32%之间。最常用的衡量水对微生物的可用性的指标是在相同温度和压力下基质中水的蒸汽压与纯水的蒸汽压之比,这个量被称为水活度 (Aw),或称为平衡相对湿度 (ERH),即ERH=Aw×100%[20]。霉菌生物膜中的水含量与空气湿度有关,较高的空气湿度有益于生物膜中保持高的水分,同时也为铝合金霉菌电化学腐蚀过程提供了良好的离子导体,水的存在是腐蚀发生的必要条件。部分霉菌生长的最低Aw值为:根霉属0.93~0.92,毛霉属0.93~0.92,黑曲霉0.88~0.80,青霉属0.83~0.80,黄曲霉0.8,白曲霉0.8,灰绿曲霉0.75~0.73,安氏曲霉0.65,整体来看霉菌对于湿度的要求还是比较高。对于大气腐蚀而言,湿热的空气可以在材料表面形成薄液膜,薄液膜条件下将会显著地加速铝合金腐蚀的阴极过程,即氧的还原,霉菌存在条件可以显著地通过生物催化作用加速氧的还原过程,从而加速铝合金材料的腐蚀尤其是局部腐蚀。
在适宜的温度和湿度以及充足的营养物质条件下,霉菌 (如图1) 具有如下特性[21-23]:(1) 霉菌体积微小,从外界环境中吸取营养和排泄废物是通过整个细胞表面进行的,具有极大的表面与体积比。极大的表面积使霉菌在短时间内吸收大量养料和排出大量代谢废物,为大量生长繁殖提供了物质基础。(2) 霉菌具有惊人的转化速度和代谢能力,在条件合适时,霉菌一昼夜合成的产物可达本身原来重量的30~40倍。霉菌的代谢越旺,则产生的腐蚀性物质越多,对铝合金腐蚀越大。(3) 霉菌由于体积小、数量多、繁殖快以及与外界的接触更为直接等原因,使得它们比较容易适应外界不良的环境。如果环境条件发生剧烈变化,部分霉菌会发生变异而适应新的环境,很难将霉菌从环境中彻底清除。从霉菌特性可看出,霉菌引发的铝合金的腐蚀过程及其复杂,影响因素众多,这也给铝合金霉菌腐蚀研究带来挑战。
2 铝合金霉菌腐蚀机制
2.1 酸蚀机制
在碳源、氮源及无机盐等营养物质充足的条件下,霉菌的孢子就会在飞机机体、油箱等区域萌芽和生长,空气中的霉菌孢子和菌丝会在被污染的铝合金结构材料表面迅速扩散,对样品的污染覆盖率超过70%[24]。有研究者[25-27]认为,霉菌在生长代谢过程中会从其吸附的铝合金表面获取一定量的金属元素 (如Fe、Cu、Zn、Mg等金属元素),可以促进铝合金部分溶解,且代谢过程中产生大量的有机酸,包括琥珀酸、草酸、戊二酸、乙醛酸、丙酮酸等,显著降低局部腐蚀介质的pH,破环铝合金表面氧化膜层,促进应力腐蚀开裂、电偶腐蚀等,两者的共同作用下可以显著地加速铝合金构件腐蚀失效。Akpan等[28]调查了尼日利亚尼日尔三角洲地区腐蚀石油管道中真菌的多样性,发现包含青霉属、曲霉菌属、角霉属等,结果表明,大部分霉菌都可以产生大量的有机酸,如甲酸、柠檬酸、乙酸和天冬氨酸等,有机酸的产生导致了管道腐蚀,是造成石油泄漏的因素之一。Wang等[29]研究认为,黑曲霉在高盐环境中可以显著地加速7075-T6铝合金均匀腐蚀和局部腐蚀,和空白相比,霉菌存在条件下,均匀腐蚀和局部腐蚀速率分别增加了3.7和22.4倍。铝合金表面霉菌生物膜形貌如图2a,可以看出生物膜包含着大量的菌丝;18 d局部腐蚀坑深度达到200多微米,如图2b和c所示。局部腐蚀是造成铝合金结构构件服役失效最为关键的因素之一,由此进一步验证了霉菌腐蚀是造成铝合金结构服役失效的重要原因之一。Wang等[29]也认为黑曲霉可以代谢产生大量的醋酸,酸蚀也是诱导铝合金局部腐蚀的原因之一。
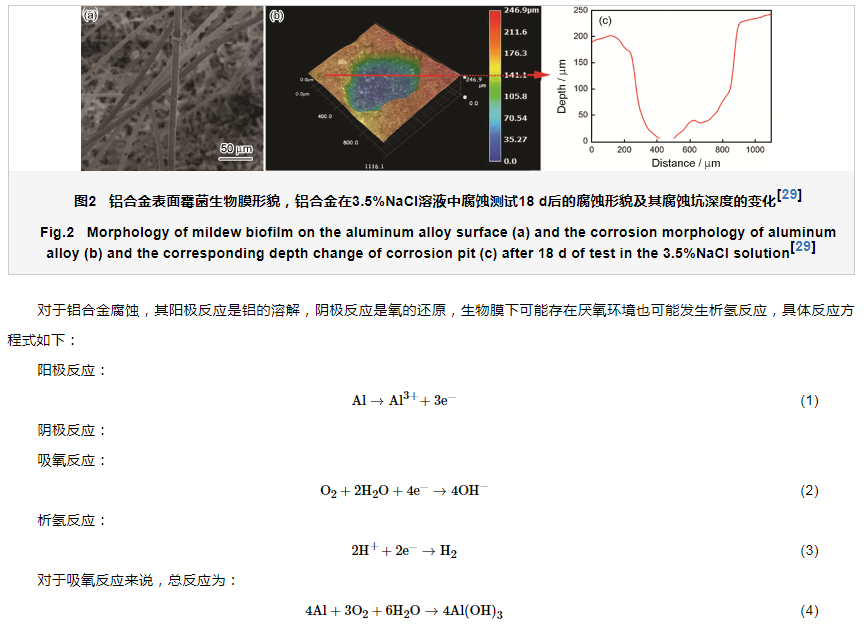
对于铝合金霉菌代谢产物腐蚀机制来说,酸蚀是导致铝合金局部腐蚀的直接原因。铝合金表面形成的生物膜极易为膜下形成一个良好的厌氧条件,而生物膜下又可以积聚大量的有机酸,从而促进阴极析氢反应,导致铝合金的局部腐蚀显著的加速。Dai等[30]对铝合金霉菌腐蚀研究表明,测试21 d后,黑曲霉产生的醋酸含量占据总的有机酸含量的96%左右,醋酸的含量在0.068 mol/L左右,微电极测试表明pH约为2.3。利用同等条件测试了醋酸溶液中铝合金腐蚀速率发现和霉菌存在条件铝合金腐蚀速率接近,从而间接地证明了霉菌的酸蚀机制。须说明的是,霉菌产生的酸的种类和产量与霉菌的种类、培养基介质以及环境条件等密切相关,铝合金霉菌酸腐蚀是多种有机酸相互协同耦合的结果。
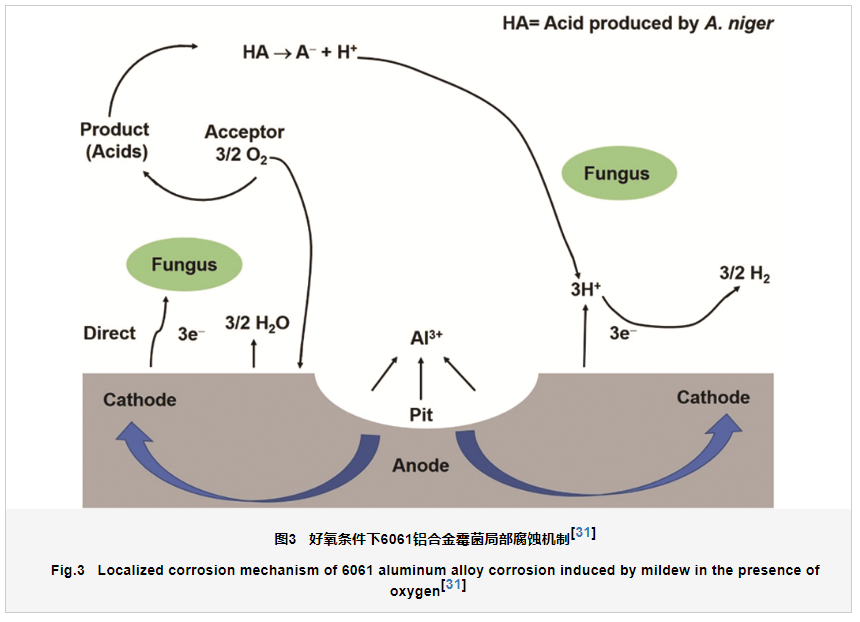
Jirón-Lazos等[31]提出了好氧条件下霉菌诱导产生的铝合金局部腐蚀机制,如图3所示。因为霉菌的新陈代谢活动以及生物膜的形成,铝合金表面处于一个微氧环境中,此时铝合金阳极溶解释放的电子一方面直接转移到霉菌体内用于氧的还原,另一方面因为霉菌代谢产生的有机酸的存在电子被析氢反应消耗掉,从而促进铝合金局部腐蚀的产生。整个腐蚀过程中,生物膜发挥着重要作用,整个腐蚀过程几乎都是在生物膜内完成的,材料表面的生物膜是非均匀的,生物膜结构和组分的差异也是导致材料局部腐蚀的关键[32]。铝合金表面霉菌生物膜具有不均一性 (图2a),从而形成大量的阳极和阴极反应活性位点,大量阳极活性位点的产生是导致铝合金发生大面积点蚀的原因。
2.2 浓差电池腐蚀机制
当同一种金属不同部位所接触的介质具有不同浓度时,引起了电极电位的不同而形成的腐蚀电池,称作浓差电池,氧浓差电池是造成金属材料局部腐蚀的主要机制之一。对于氧浓差电池,富氧区电极电位较正,为金属阴极;而贫氧区电极电位较负,为金属阳极。氧浓差电池的形成可以显著地加速金属材料阳极区的溶解,从而导致局部腐蚀[33,34]。海洋环境中,铝合金典型的局部腐蚀机制如图4所示[35]。从图4可以看出,在无霉菌存在条件下,Cl-存在下会破坏铝合金表面钝化膜,进而诱发点蚀,随着腐蚀的进行,腐蚀产物Al(OH)3在点蚀坑部位的堆积直接造成了腐蚀坑内的厌氧环境,进而形成氧浓差电池。在氯离子催化作用下,腐蚀坑内作为阳极发生氧化反应 (反应1) 释放电子,电子传递到其他部位用于氧气的还原 (反应2),从而显著地加速铝合金局部腐蚀,导致铝合金材料失效。在霉菌存在条件,铝合金氧浓差腐蚀机制更为复杂。霉菌在铝合金表面吸附之后很快就会形成一层生物膜,生物膜一旦形成之后很难彻底剥离和去除,霉菌腐蚀过程与其在材料表面形成的生物膜密切相关。生物膜主要是由于微生物在材料表面特定的活性位点上附着,进一步在微生物自身生命活动及其与基体材料作用下形成一种复杂凝胶状混合物,主要成分是水、腐蚀产物和胞外聚合物 (EPS)。EPS的主要成分是多糖、蛋白质、核酸和脂类。生物膜的形成是伴随着微生物的生长和消亡以及环境不断变化的动态过程[36]。因此,铝合金霉菌腐蚀也是一个动态的过程。
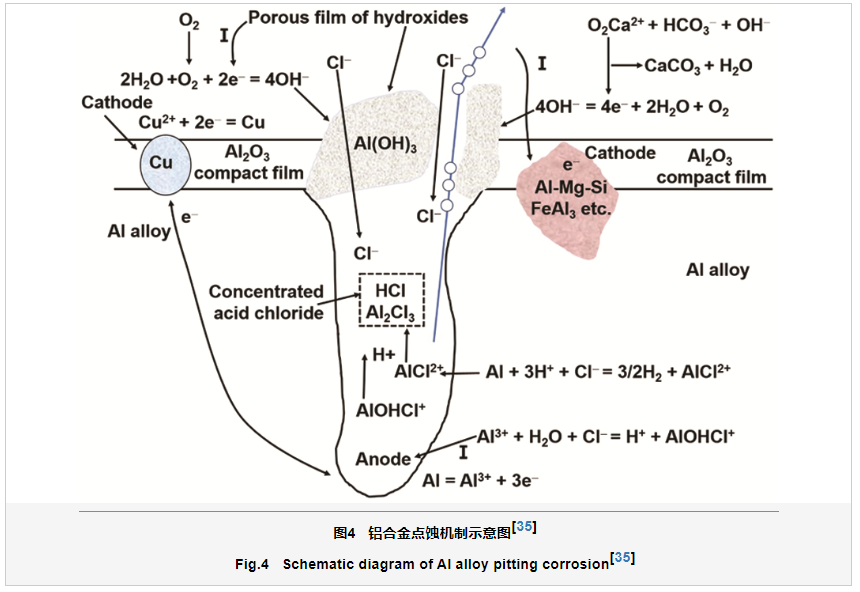
霉菌是好氧型微生物,其生长代谢过程中会消耗大量的氧气。生物膜形成后期,在生物膜下因为霉菌对氧气的大量消耗会伴随着产生厌氧和缺氧区,另外腐蚀产物局部堆积也会限制氧的扩散从而也导致厌氧区的产生[37]。Imo等[38]研究了烟曲霉菌对铝的腐蚀行为,结果发现,铝腐蚀速率增加是由生物膜的不均一性导致的氧浓差电池引起的。霉菌生物膜下氧浓差电池腐蚀机制更为复杂,首先生物膜的附着和形成可以导致更多贫氧区的产生,从而为氧浓差电池的形成创造了良好的条件;其次,霉菌在代谢过程中会代谢大量的有机酸,对于铝合金表面氧化膜的破坏性增强,可以直接促进形成更多点蚀活性位点;另外,初期酸蚀可能是导致点蚀快速生长的一个重要原因。这些因素都会对图4中铝合金点蚀的生长和扩展产生重要影响,但是近年来关于铝合金霉菌作用的氧浓差腐蚀机制报道并不多见,后续研究仍然需要加强。
2.3 其他腐蚀机制
Silva等[39]研究发现,黑曲霉可以改变本体溶液的pH值,通过原子力显微镜测试表明,黑曲霉在铝合金表面吸附的部位发生了明显的腐蚀,这表明霉菌可以直接与铝合金相互作用,影响铝合金的腐蚀过程,但是霉菌是如何通过直接接触改变铝合金腐蚀行为目前尚不清楚。Corvo等[26]研究发现,和阳极氧化铝合金材料相比,整体而言非阳极氧化铝合金表面霉菌的量要高出一个数量级左右,这也说明霉菌更倾向于在铝合金表面生长,而且霉菌能够与铝合金直接相互作用,一方面促进霉菌的生长代谢过程,另一方面也加速了铝合金的腐蚀破坏。其他研究者也确认了铝合金表面处理差异与其表面霉菌菌量之间的上述对应关系[31]。
Jirón-Lazos等 [31]提出了霉菌和铝合金直接电子传递 (DET) 机制,揭示了霉菌直接电子传递和酸蚀耦合局部腐蚀机制 (图3),但是需要说明的是,霉菌和铝合金界面电子直接传递机制目前尚缺乏直接的证据证明。先前的报道表明,霉菌存在条件下,铝合金的阴极和阳极反应速度都明显增加[29]。对于阳极反应铝的溶解,霉菌的存在很有可能是由于加速了电子的转移从而促进了铝的阳极过程,在这个过程中,霉菌是非常有可能通过直接转移电子途径加速铝的溶解。上述霉菌和铝合金的直接相互作用也可以间接地证明[26]。对于铝合金腐蚀的阴极反应可能涉及到吸氧和析氢反应,析氢反应主要来自霉菌代谢产生的有机酸,上述已经详细说明,在此不赘述。然而对于吸氧反应来说,由于霉菌是好氧微生物,其通过自身生物催化可以显著地加速氧的还原,阴极反应的加速反过来也可以促进电子的转移,导致阳极反应速度的增加[40]。先前的研究工作中已经证明厌氧的硫酸盐还原菌 (SRB) 在缺少碳源的条件下能够在细胞表面产生纳米线从而通过直接电子传递从铁基体获得电子生长,直接加速碳钢的局部腐蚀[40,41]。但是对于霉菌来说,目前尚没有发现霉菌能够在特定环境和条件下在细胞表面产生纳米线用于获取铝合金释放的电子。目前研究发现,微生物腐蚀过程中电子传递途径主要包括[36]:(1) 细胞壁直接与金属表面接触:电子通过细胞色素C (Cytochrome C) 等膜蛋白进行传递;(2) 依靠生物纳米线的电子转移:细菌通过形成生物纳米线 (pili) 与金属形成电子传递的桥梁;(3) 电子载体依赖型的电子转移:电子通过电子载体如核黄素和黄素腺嘌呤二核苷酸 (FAD) 进入细胞膜。
目前铝合金霉菌腐蚀研究主要以腐蚀行为为主,霉菌和铝合金之间的电子转移过程研究仍然比较匮乏。因此后续研究中,非常有必要通过原位和非原位的手段来着手研究霉菌腐蚀电子转移路径及影响因素,从而为铝合金霉菌腐蚀防护提供理论依据和指导。
在海洋环境中,铝合金构件表面Cl-是无法避免的,Cl-的存在会对铝合金表面钝化膜产生严重的破坏作用,从而诱发铝合金局部腐蚀。先前的报道表明,霉菌和Cl-可以通过协同耦合作用加速铝合金的腐蚀过程 [30]。在铝合金局部腐蚀破坏过程中,Cl-主要起到自催化的作用,加速Al的溶解,如图4。霉菌在该过程中既可以起到生物的催化作用,又可以参与到铝合金腐蚀的阴阳极反应中,如上所述。因此,在实际环境中,铝合金的破坏过程和腐蚀机制更为复杂,不能单一的归结到某一因素中,而是一个多因素耦合的过程。但是在整个铝合金腐蚀破坏的过程中,研究结果表明,微生物包括霉菌在内起到非常重要的作用。近年来微生物腐蚀机制研究已经取得了很大的进展,在各种环境中的硫酸盐还原菌腐蚀机制已经逐渐清晰,但是微生物的种类和数量巨大,微生物腐蚀机制非常复杂,实验室条件又很难真实地模拟自然环境。因此,对于铝合金霉菌腐蚀研究来说,未来仍然需要更多的科研工作者进行大量的研究工作,借助目前现有的先进技术,从微观尺度原位研究霉菌腐蚀行为、电子转移路径是非常有必要的,但是也面临着巨大的挑战。
3 铝合金霉菌腐蚀防护
微生物腐蚀控制策略目前普遍采用的方法主要有杀菌剂、缓蚀剂、抗菌涂层、生物抑制法等,杀菌剂是目前应用最为广泛的控制微生物腐蚀的方法,但是杀菌剂大规模的使用会对环境造成二次污染[42-44]。作者在先前的工作中尝试利用物理磁场来控制微生物腐蚀,且取得了很不错的效果[45]。对于铝合金霉菌腐蚀来说,其防护策略可以从两个方面着手。第一,对于封闭和半封闭环境中,使用杀菌剂可以有效地控制霉菌腐蚀,但是因为生物膜的保护作用,杀菌剂对于生物膜中的霉菌的杀灭效果不是很理想[46]。第二,可以通过构筑抗菌涂层及其他特性涂层防止霉菌在铝合金构件表面的附着,从而起到控制霉菌腐蚀的目的[47,48]。
目前关于铝合金霉菌腐蚀控制方法研究的报道仍然有限。Wang等[49]研究了环境友好型和高效杀菌剂吡啶硫酮锌对铝合金霉菌腐蚀的影响,结果表明,吡啶硫酮锌使用浓度仅为20 mg/L时,可以显著地抑制霉菌在铝合金表面的附着和吸附以及生物膜的形成,铝合金腐蚀速率下降了94%。Imo等[50]研究认为,胡椒提取物可以有效抑制烟曲霉的生长,从而起到控制铝合金霉菌腐蚀目的,提取物的最低霉菌抑制浓度在10~19.5 mg/L之间,与提取条件有关。石娇等[51]在铝合金表面构筑了一种有机硅改性环氧树脂复合涂料,结果表明,该涂层能够显著提高铝合金耐霉菌腐蚀性能,其加入绢云母粉则效果更好。赵立华等[52]研究认为,高强铝合金阳极氧化层耐霉性能良好,炮油涂层、涂漆层耐霉性能较差。
铝合金构件表面一旦形成霉菌生物膜,将很难彻底清除[53]。因此,未来铝合金霉菌防护领域研究首先要着重开发先进环境友好型抗菌涂层,从源头上控制霉菌在铝合金构件材料表面的吸附。另外在已经形成霉菌生物膜的构件表面,可以使用一些霉菌杀菌剂直接将生物膜中霉菌杀死,但是霉菌在特定条件下会通过基因变异而产生抗药性;也可以采取一定的措施将霉菌生物膜彻底清除,但是霉菌生物膜的完全去除仍然面临着巨大的挑战。
4 总结和展望
沿海湿热环境,霉菌腐蚀问题严重威胁着各种高端装备铝合金构件的长期安全服役,铝合金霉菌腐蚀机制和防护研究已经取得了一定的进展,但是铝合金的霉菌腐蚀问题仍然还没有得到彻底解决。首先,霉菌的种类较多,对铝合金具有腐蚀性的霉菌的分离和鉴定仍然是研究重点之一,霉菌和铝合金之间的相互作用关系及生物膜形成的关键影响因素也有待进一步阐明;其次,铝合金的霉菌腐蚀机制,尤其是腐蚀过程中电子传递机制以及霉菌在铝合金腐蚀过程中是如何发挥作用的仍然需要进一步研究,霉菌除酸以外其他代谢产物对铝合金腐蚀过程的影响仍然不明;最后,铝合金霉菌腐蚀的监检测和防护研究,是解决铝合金霉菌腐蚀问题的关键。
未来铝合金霉菌腐蚀与防护研究中,第一要借助当前先进的分子生物学技术,对湿热海洋环境中霉菌的种类进行分离鉴定,建立霉菌种类大数据库,为霉菌腐蚀研究创造基础;第二,要设计合理的原位测试装置,结合实验室和现场检测,原位研究铝合金霉菌腐蚀过程,着重从微观尺度通过直接和间接的方法揭示铝合金霉菌腐蚀过程中的电子转移路径以及霉菌和铝合金之间相互作用关系,为铝合金霉菌腐蚀防护提供理论依据;第三,要设计合理的原位对霉菌响应的监检测手段,实时掌握铝合金构件表面霉菌附着情况;第四,结合目前现有的铝合金防霉技术和方法,着重开发低毒、高效、环保的防霉杀菌剂,设计适用于海洋环境的多功能抗霉菌附着涂层,为解决铝合金霉菌腐蚀问题提供新的思路和方法。
参考文献
[1]
Reboul M C, Baroux B.
Metallurgical aspects of corrosion resistance of aluminium alloys
[J]. Mater. Corros., 2011, 62: 215
[本文引用: 1]
[2]
Arrabal R, Mingo B, Pardo A, et al.
Pitting corrosion of rheocast A356 aluminium alloy in 3.5wt.%NaCl solution
[J]. Corros. Sci., 2013, 73: 342
[本文引用: 1]
[3]
Dursun T, Soutis C.
Recent developments in advanced aircraft aluminium alloys
[J]. Mater. Des., 2014, 56: 862
[本文引用: 1]
[4]
Ma T, Wang Z Y, Han W.
A review of atmospheric corrosion of aluminum and aluminum alloys
[J]. Corros. Sci. Prot. Technol., 2004, 16: 155
[本文引用: 2]
马腾, 王振尧, 韩薇。
铝和铝合金的大气腐蚀
[J]. 腐蚀科学与防护技术, 2004, 16: 155
[本文引用: 2]
[5]
Venugopal A, Panda R, Manwatkar S, et al.
Effect of micro arc oxidation treatment on localized corrosion behaviour of AA7075 aluminum alloy in 3.5%NaCl solution
[J]. Trans. Nonferrous Met. Soc. China, 2012, 22: 700
[本文引用: 1]
[6]
Jia R, Tan J L, Jin P, et al.
Effects of biogenic H2S on the microbiologically influenced corrosion of C1018 carbon steel by sulfate reducing Desulfovibrio vulgaris biofilm
[J]. Corros. Sci., 2018, 130: 1
[本文引用: 1]
[7]
Li Y C, Xu D K, Chen C F, et al.
Anaerobic microbiologically influenced corrosion mechanisms interpreted using bioenergetics and bioelectrochemistry: a review
[J]. J. Mater. Sci. Technol., 2018, 34: 1713
[8]
Liu T, Pan S, Wang Y N, et al.
Pseudoalteromonas lipolytica accelerated corrosion of low alloy steel by the endogenous electron mediator pyomelanin
[J]. Corros. Sci., 2020, 162: 108215
[本文引用: 1]
[9]
Jia R, Unsal T, Xu D K, et al.
Microbiologically influenced corrosion and current mitigation strategies: A state of the art review
[J]. Int. Biodeter. Biodegr., 2019, 137: 42
[本文引用: 1]
[10]
Liu H W, Gu T Y, Zhang G A, et al.
Corrosion of X80 pipeline steel under sulfate-reducing bacterium biofilms in simulated CO2-saturated oilfield produced water with carbon source starvation
[J]. Corros. Sci., 2018, 136: 47
[本文引用: 1]
[11]
Emerson D.
The role of iron-oxidizing bacteria in biocorrosion: A review
[J]. Biofouling, 2018, 34: 989
[12]
Liu T, Wang Y N, Pan S, et al.
The addition of copper accelerates the corrosion of steel via impeding biomineralized film formation of Bacillus subtilis in seawater
[J]. Corros. Sci., 2019, 149: 153
[本文引用: 1]
[13]
Wang X H, Wang L.
A study of measures and test technology for fungus resistance to weaponry
[J]. Aeronaut. Stand. Qual., 2003, (2): 38
[本文引用: 1]
王晓慧, 王丽。
武器装备防霉措施和试验技术探讨
[J]. 航空标准化与质量, 2003, (2): 38
[本文引用: 1]
[14]
Qu X Y, Deng L.
Analysis of the environmental worthiness of shipborne weapons in marine environment
[J]. Ship Electron. Eng., 2011, 31(4): 138
[本文引用: 1]
曲晓燕, 邓力。
舰载武器海洋环境适应性分析
[J]. 舰船电子工程, 2011, 31(4): 138
[本文引用: 1]
[15]
McNamara C J, Perry IV T D, Leard R, et al.
Corrosion of aluminum alloy 2024 by microorganisms isolated from aircraft fuel tanks
[J]. Biofouling, 2005, 21: 257
[本文引用: 1]
[16]
Hagenauer A, Hilpert R, Hack T.
Microbiological investigations of corrosion damages in aircraft
[J]. Mater. Corros., 1994, 45: 355
[本文引用: 1]
[17]
Li X, Wang X H.
Overview of three-proof design on carrier-based aircraft
[J]. Equip. Environ. Eng., 2006, 3(4): 12
[本文引用: 1]
李星, 王晓慧。
舰载机三防设计技术研究综述
[J]. 装备环境工程, 2006, 3(4): 12
[本文引用: 1]
[18]
Cheng P, Song W, Chen L, et al.
Molecular characterization of Aspergillus tubingensis and Eurotium amstelodami associated with black brick tea
[J]. Int. J. Agric. Biol., 2016, 18: 489
[本文引用: 3]
[19]
Zhang J T, Wu L Y.
Mildew protection design of airborne equipments
[J]. Equip. Environ. Eng., 2007, 4(6): 70
[本文引用: 1]
张江涛, 吴龙益。
机载设备霉菌防护设计
[J]. 装备环境工程, 2007, 4(6): 70
[本文引用: 1]
[20]
Ayerst G.
The effects of moisture and temperature on growth and spore germination in some fungi
[J]. J. Stored Prod. Res., 1969, 5: 127
[本文引用: 1]
[21]
Martin-Sanchez P M, Gorbushina A A, Kunte H J, et al.
A novel qPCR protocol for the specific detection and quantification of the fuel-deteriorating fungus Hormoconis resinae
[J]. Biofouling, 2016, 32: 635
[本文引用: 1]
[22]
Zhu W F, Li K, Wang B, et al.
Practical fungus-proof technology and experiment research of carrier-based aircraft hydraulic system
[J]. Hydraul. Pneumat. Seals, 2014, 34(1): 71
朱武峰, 李昆, 王兵等。
舰载机液压系统实用防霉技术及试验研究
[J]. 液压气动与密封, 2014, 34(1): 71
[23]
Qu Q, Wang L, Li L, et al.
Effect of the fungus, Aspergillus niger, on the corrosion behaviour of AZ31B magnesium alloy in artificial seawater
[J]. Corros. Sci., 2015, 98: 249
[本文引用: 1]
[24]
Zhou W, Zhang Z G.
Analysis on corrosion effect of mold pollution on aviation aluminum alloy equipment
[J]. Environ. Sci. Manage., 2019, 44(1): 87
[本文引用: 1]
周伟, 张作刚。
霉菌污染对航空铝合金设备腐蚀作用分析研究
[J]. 环境科学与管理, 2019, 44(1): 87
[本文引用: 1]
[25]
Mie?inskas P, Leinartas K, Uksien? V, et al.
QCM study of microbiological activity during long-term exposure to atmosphere—aluminium colonisation by Aspergillus Niger
[J]. J. Solid State Electrochem., 2007, 11: 909
[本文引用: 1]
[26]
Corvo F, Jirón-Lazos U, de la Rosa S, et al.
Aluminum and anodized aluminum biocorrosion caused by Aspergillus niger
[R]. Mexico, 2016: 9964
[本文引用: 2]
[27]
De Leo F, Campanella G, Proverbio E, et al.
Laboratory tests of fungal biocorrosion of unbonded lubricated post-tensioned tendons
[J]. Constr. Build. Mater., 2013, 49: 821
[本文引用: 1]
[28]
Akpan G U, Iliyasu M.
Fungal populations inhabiting biofilms of corroded oil pipelines in the Niger Delta region of Nigeria
[J]. Sky J. Microbiol. Res., 2015, 3: 36
[本文引用: 1]
[29]
Wang J L, Xiong F P, Liu H W, et al.
Study of the corrosion behavior of Aspergillus niger on 7075-T6 aluminum alloy in a high salinity environment
[J]. Bioelectrochemistry, 2019, 129: 10
[本文引用: 5]
[30]
Dai X Y, Wang H, Ju L K, et al.
Corrosion of aluminum alloy 2024 caused by Aspergillus niger
[J]. Int. Biodeter. Biodegr., 2016, 115: 1
[本文引用: 2]
[31]
Jirón-Lazos U, Corvo F, De la Rosa S C, et al.
Localized corrosion of aluminum alloy 6061 in the presence of Aspergillus niger
[J]. Int. Biodeter. Biodegr., 2018, 133: 17
[本文引用: 5]
[32]
Dong Z H, Shi W, Ruan H M, et al.
Heterogeneous corrosion of mild steel under SRB-biofilm characterised by electrochemical mapping technique
[J]. Corros. Sci., 2011, 53: 2978
[本文引用: 1]
[33]
Machuca L L, Bailey S I, Gubner R, et al.
Effect of oxygen and biofilms on crevice corrosion of UNS S31803 and UNS N08825 in natural seawater
[J]. Corros. Sci., 2013, 67: 242
[本文引用: 1]
[34]
Heyer A, D'Souza F, Morales C F L, et al.
Ship ballast tanks a review from microbial corrosion and electrochemical point of view
[J]. Ocean Eng., 2013, 70: 188
[本文引用: 1]
[35]
Liu S Y.
Preparation of multifunctional anti-corrosion polyaniline composite coating on Al alloy under marine environment
[D]. Hefei: University of Science and Technology of China, 2019
[本文引用: 3]
刘素云。
海洋环境用Al合金表面多功能耐蚀聚苯胺复合涂层的研制
[D]. 合肥: 中国科学技术大学, 2019
[本文引用: 3]
[36]
Liu H W, Xu D K, Wu Y N, et al.
Research progress in corrosion of steels induced by sulfate reducing bacteria
[J]. Corros. Sci. Prot. Technol., 2015, 27: 409
[本文引用: 2]
刘宏伟, 徐大可, 吴亚楠等。
微生物生物膜下的钢铁材料腐蚀研究进展
[J]. 腐蚀科学与防护技术, 2015, 27: 409
[本文引用: 2]
[37]
Qu Q, Li S L, Li L, et al.
Adsorption and corrosion behaviour of Trichoderma harzianum for AZ31B magnesium alloy in artificial seawater
[J]. Corros. Sci., 2017, 118: 12
[本文引用: 1]
[38]
Imo E O, Orji J C, Nweke C O.
Influence of Aspergillus fumigatus on corrosion behaviour of mild steel and aluminium
[J]. Int. J.
Appl
Microbiol. Biotechnol. Res., 2018, 6: 61
[本文引用: 1]
[39]
Silva A M A, Santiago T M, Alves C R, et al.
An evaluation of the corrosion behavior of aluminum surfaces in presence of fungi using atomic force microscopy and other tests
[J]. Anti-Corros. Methods Mater., 2007, 54: 289
[本文引用: 1]
[40]
Rosales B M, Iannuzzi M.
Aluminium AA2024 T351 aeronautical alloy: Part 1. Microbial influenced corrosion analysis
[J]. Mater. Sci. Eng., 2008, A472: 15
[本文引用: 2]
[41]
Dou W W, Liu J L, Cai W Z, et al.
Electrochemical investigation of increased carbon steel corrosion via extracellular electron transfer by a sulfate reducing bacterium under carbon source starvation
[J]. Corros. Sci., 2019, 150: 258
[本文引用: 1]
[42]
Liu H W, Gu T Y, Lv Y L, et al.
Corrosion inhibition and anti-bacterial efficacy of benzalkonium chloride in artificial CO2-saturated oilfield produced water
[J]. Corros. Sci., 2017, 117: 24
[本文引用: 1]
[43]
Ma F L, Li J L, Zeng Z X, et al.
Tribocorrosion behavior in artificial seawater and anti-microbiologically influenced corrosion properties of TiSiN-Cu coating on F690 steel
[J]. J. Mater. Sci. Technol., 2019, 35: 448
[44]
Jia R, Yang D Q, Rahman H B A, et al.
Laboratory testing of enhanced biocide mitigation of an oilfield biofilm and its microbiologically influenced corrosion of carbon steel in the presence of oilfield chemicals
[J]. Int. Biodeter. Biodegr., 2017, 125: 116
[本文引用: 1]
[45]
Liu H W, Gu T Y, Zhang G A, et al.
The effect of magneticfield on biomineralization and corrosion behavior of carbon steel induced by iron-oxidizing bacteria
[J]. Corros. Sci., 2016, 102: 93
[本文引用: 1]
[46]
Arafat El S S, Matzdorf C A, Spadafora S J, et al.
Composition and process for removing and preventing mildew and fungal growth
[P]. USA Pat, 7494670, 2009
[本文引用: 1]
[47]
Akid R, Wang H, Smith T J, et al.
Biological functionalization of a Sol-Gel coating for the mitigation of microbial‐induced corrosion
[J]. Adv. Funct. Mater., 2008, 18: 203
[本文引用: 1]
[48]
Liu Q Q, Lu L, Gao G, et al.
Research progress on fungal corrosion of metals and their protective layers in atmos-pheric environments
[J]. Chin. J. Eng., 2017, 39: 1463
[本文引用: 1]
刘倩倩, 卢琳, 高歌等。
大气环境中金属及其保护层霉菌腐蚀研究的进展
[J]. 工程科学学报, 2017, 39: 1463
[本文引用: 1]
[49]
Wang J L, Li C J, Zhang X X, et al.
Corrosion behavior of Aspergillus niger on 7075 aluminum alloy and the inhibition effect of zinc pyrithione biocide
[J]. J. Electrochem. Soc., 2019, 166: G39
[本文引用: 1]
[50]
Imo E O, Orji J C, Nweke C O.
Corrosion and fungal growth inhibiting effects of Piper guineense extracts
[J]. J. Appl. Environ. Microbiol., 2018, 6: 37
[本文引用: 1]
[51]
Shi J, Qu Y P.
Study on organic coatings for mould corrosion resistance in ocean climate
[J]. Surf. Technol., 2011, 40(1): 56
[本文引用: 1]
石娇, 曲彦平。
耐海洋环境中霉菌腐蚀有机涂层的研究
[J]. 表面技术, 2011, 40(1): 56
[本文引用: 1]
[52]
Zhao L H, Duan Y P.
Mould resistance of protective layer on high-strength alloy structural steel and high-strength aluminum alloy
[J]. Equip. Environ. Eng., 2015, 12(4): 82
[本文引用: 1]
赵立华, 段渝平。
高强度合金结构钢与高强度铝合金防护层的耐霉性研究 [J]. 装备环境工程, 2015, 12(4): 82 [本文引用: 1]
[53] Brown T T, Lee J S. Microscopic evaluation of fungal cleaning protocols for aircraft coatings [J]. Microsc. Microanal., 2019, 25 (): 718 [本文引用: 1]
免责声明:本网站所转载的文字、图片与视频资料版权归原创作者所有,如果涉及侵权,请第一时间联系本网删除。
相关文章

官方微信
《腐蚀与防护网电子期刊》征订启事
- 投稿联系:编辑部
- 电话:010-62316606-806
- 邮箱:fsfhzy666@163.com
- 腐蚀与防护网官方QQ群:140808414
点击排行
PPT新闻

“海洋金属”——钛合金在舰船的
点击数:8148

腐蚀与“海上丝绸之路”
点击数:6486