
抗菌肽在海洋防污领域的研究进展
2022-07-28 14:12:11
作者:楼彤,白秀琴,贺小燕,杨膺杨宗澄,袁成清 来源:中国表面工程
分享至:


摘要
海洋生物污损仍是目前急需解决的难题,利用抗菌肽对基体进行改性获得防污效果是一种有效的绿色应对策略,然而目前缺少抗菌肽在海洋防污领域应用的全面报道。基于抗菌肽的二级结构,在详细介绍 α-螺旋结构、β-折叠结构和延伸/弹性结构三类不同结构抗菌肽的基础上,总结抗菌肽引发的膜损伤以及非膜损伤的多种抗菌机理,明晰抗菌肽的作用靶点:细胞膜、酶、蛋白质和核酸;阐述几种目前常见的基体表面接枝抗菌肽的方法;基于常见的海洋细菌、硅藻和大型污损生物繁殖体等典型海洋污损生物,对抗菌肽的防污特性进行分析。最后提出抗菌肽的制备、表面接枝方法,以及构效关系是其在海洋防污领域应用所面临的难题与未来发展方向,填补了抗菌肽防污技术领域目前缺少此类综述的空白。
关键词: 生物污损 ; 抗菌肽 ; 抗菌机理 ; 接枝方法 ; 防污性能
0 前言
我国是世界贸易大国,海运承担了大部分的进出口货物运输,随着“21世纪海上丝绸之路”战略构想的提出,海运在沿线国家和地区间经贸往来和文化交流中的重要桥梁和纽带作用更加凸显。2014年国务院印发《关于促进海运业健康发展的若干意见》,将海运业发展上升为国家战略。2020年末,远洋运输船舶达到1 499艘,运力达到180.8万标准箱。船舶作为海运的重要载体,在海运业中起着大动脉的作用,快速、高效、节能、环保已成为其发展目标。然而,海洋生物在船舶表面的附着会造成严重的生物污损现象。海洋生物污损是指海洋生物在基体表面非预期的附着。生物污损会造成许多问题,尤其是在船舶上。污损生物在船舶浸水部位的附着,导致船舶航行时摩擦阻力剧增,燃油消耗增加,维护成本增大。同时,生物污损使得几乎所有船舶浸水部位的减阻技术无法在实际工程中得到有效应用[1]。因此,船舶生物污损已成为海运业面临的重大技术难题,解决生物污损以降低船舶燃油消耗,对实现船舶低能耗、高效率运行以及促进海运业的高质量发展具有重要意义。
国内外科研工作者从化学、物理和生物三方面入手构建了多种防污方法来对抗海洋生物污损。化学防污方法主要是利用化学物质对海洋污损生物的毒杀作用,杀死海洋污损生物或抑制其附着。传统的化学防污方法主要包括有机锡类防污涂料和含铜、锌、硅等无锡自抛光防污涂料[2-3]。然而由于环境保护的严格要求,此类防污涂料终会被禁用,开发出可替代的绿色防污涂料迫在眉睫。华南理工大学张广照团队采用生物降解高分子防污材料替代自抛光防污涂料中的毒性物质,开发了新型环境友好的化学防污涂料[4]。物理防污方法是利用材料表面的物理性质获得防污功能的一类环境友好型防污材料,主要有表面微结构涂层、低表面能涂层、表面亲/疏水涂层、表面植绒涂层、导电涂层以及液体灌注多孔表面[3]。生物防污方法主要是通过提取生物体内的活性物质,来抑制污损生物的粘附。提取物的种类多样,包括生物肽、生物酶、生物碱、菇类化合物和类固醇等[5]。在众多防污方法中,利用防污剂防污是目前最方便、有效和经济的方法。海洋生物给绿色无毒防污剂的来源提供了极大的选择空间。在自然界中,动植物可以诱导产生一类具有抗菌活性的多肽物质,称抗菌肽(AMPs)。抗菌肽在海洋生物体内普遍存在,因其具有广谱抗菌活性、低浓度快速杀菌能力和环境友好性而成为防污领域的热点。例如,雪蟹通过分泌一种抗菌肽来保护自身免受有害细菌的入侵,以保持自身清洁[6]。短沟对虾中提取的抗菌肽Crustin具有杀菌活性,同时可以抑制生物膜的形成[7]。近年来,抗菌肽在海洋防污领域的应用也取得了一定的进展[5],然而缺少目前发展现状的全面总结。本文从抗菌肽的分类、抗菌机理、基体表面接枝方法以及海洋防污性能等方面,阐述目前抗菌肽在防污领域的研究进展,并提出了未来的发展方向和所面临的难题,以期为后续抗菌肽防污技术的发展和应用提供参考。
1 抗菌肽的分类
自从20世纪80年代,昆虫天蚕素、人类防御素、蛙皮素等性能优异的抗菌肽被发现以来,抗菌肽的家族在不断扩大,抗菌肽数据库(Antimicrobial peptide database,APD)记录了来自六大类生物(细菌、古细菌、原生生物、真菌、植物、动物)的3 000多种抗菌肽,其中动物占75%[8]。抗菌肽稳定的二级结构是其杀菌活性的来源,从而可以影响其防污活性。通常依靠其已知的结构,如 α-螺旋结构、β-折叠结构和延伸/弹性结构对抗菌肽进行分类(图1)。
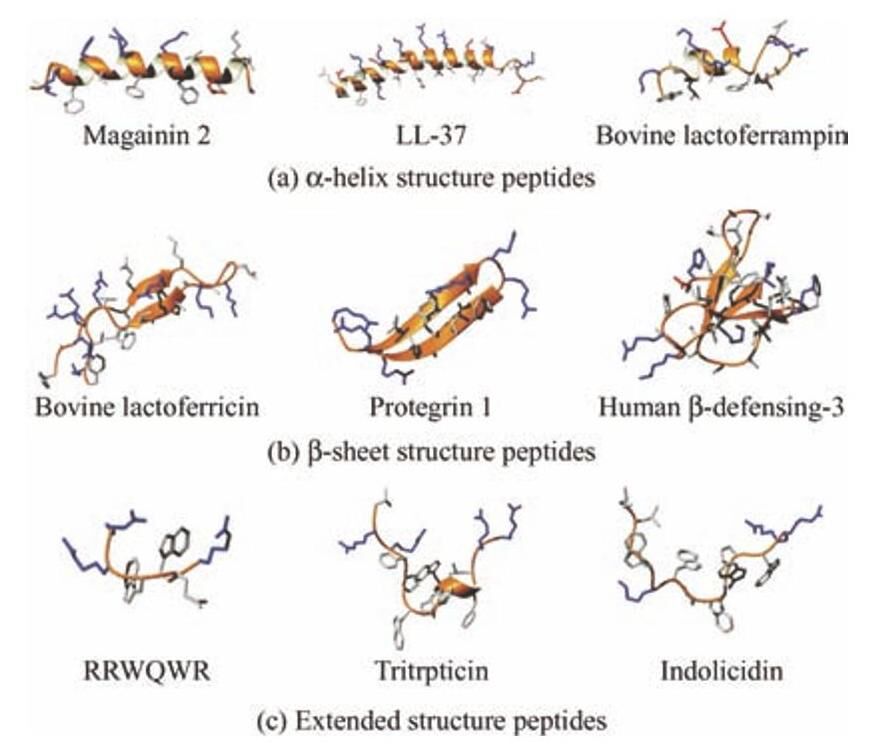
图1 抗菌肽的二级结构[16]
1.1 α-螺旋结构抗菌肽
α-螺旋结构肽的骨架围绕着一个中心轴以右手螺旋的方式规律地伸展,每隔3.6个残基螺旋上升一圈。由于形成 α-螺旋结构所带来的肽主链上氢键的焓增益不能抵消维持该受限构象的熵代价,该类肽在海水等溶液环境中没有呈现出明显的 α-螺旋结构,需要一个膜环境来诱导进行适当的折叠,才会形成 α-螺旋结构[9]。这种结构的转变导致了亲水残基与疏水残基在空间上沿 α-螺旋轴分隔,使其拥有两亲性[10]。两亲性的形成有利于肽在初期与脂质膜的结合,带电荷的一面朝向膜的磷脂头基从而平铺在表面上。尾部的性质会影响肽插入膜的深度,进而影响其抗菌活性。
1.2 β-折叠结构抗菌肽
β-折叠结构肽的骨架呈锯齿状的方式按层规律地排列,两残基间距为0.65nm(顺式)或0.70nm (反式)。该类肽也需要经过膜环境的诱导才能形成 β-折叠结构[11]。β-折叠结构肽包含2~10个半胱氨酸,从而形成1~5个二硫键来稳定其生物活性构象。这些二硫键不会影响抗菌肽的抗菌活性,速普肽和乳铁蛋白肽的线性衍生物同样保持了抗菌活性[12-13]。防御素就是其中一类在特定位置保存有6~8个半胱氨酸的 β-折叠结构肽,它在天然免疫系统中发挥重要作用,能有效杀死多种微生物。
1.3 延伸/弹性结构抗菌肽
高比例脯氨酸和甘氨酸残基的存在通常会破坏抗菌肽的典型二级结构,使其变成延伸/弹性结构[14]。大部分富含脯氨酸的抗菌肽是从哺乳动物 (如indolicidin、tritrpticin)和昆虫(如apidaecins) 中分离出来的。许多延伸/弹性肽不具有膜活性,而是作用于细胞内部物质[15]。
2 抗菌机理
低浓度的抗菌肽拥有高效的抗菌活性,主要通过破坏完整的膜结构或抑制细胞的某些关键过程来影响细菌的生长繁殖,如图2所示。尽管不同的抗菌肽序列长短差别很大,抗菌机制也有所不同,但都必须经过一定的步骤来杀死细菌:首先,肽通过静电作用靠近细菌;然后,肽穿过细菌细胞壁的屏障与细胞质膜上的磷脂双分子层相互作用,平行地在磷脂双分子层表面聚集,随着浓度的增加,肽开始垂直于膜方向;最后,当达到活性浓度阈值时(一般肽脂摩尔比为1∶20 [17]),肽分子开始发挥其抗菌功能,直接作用于细胞膜造成膜损伤或者转移到细胞质中,通过非膜损伤的方式来杀死细菌。
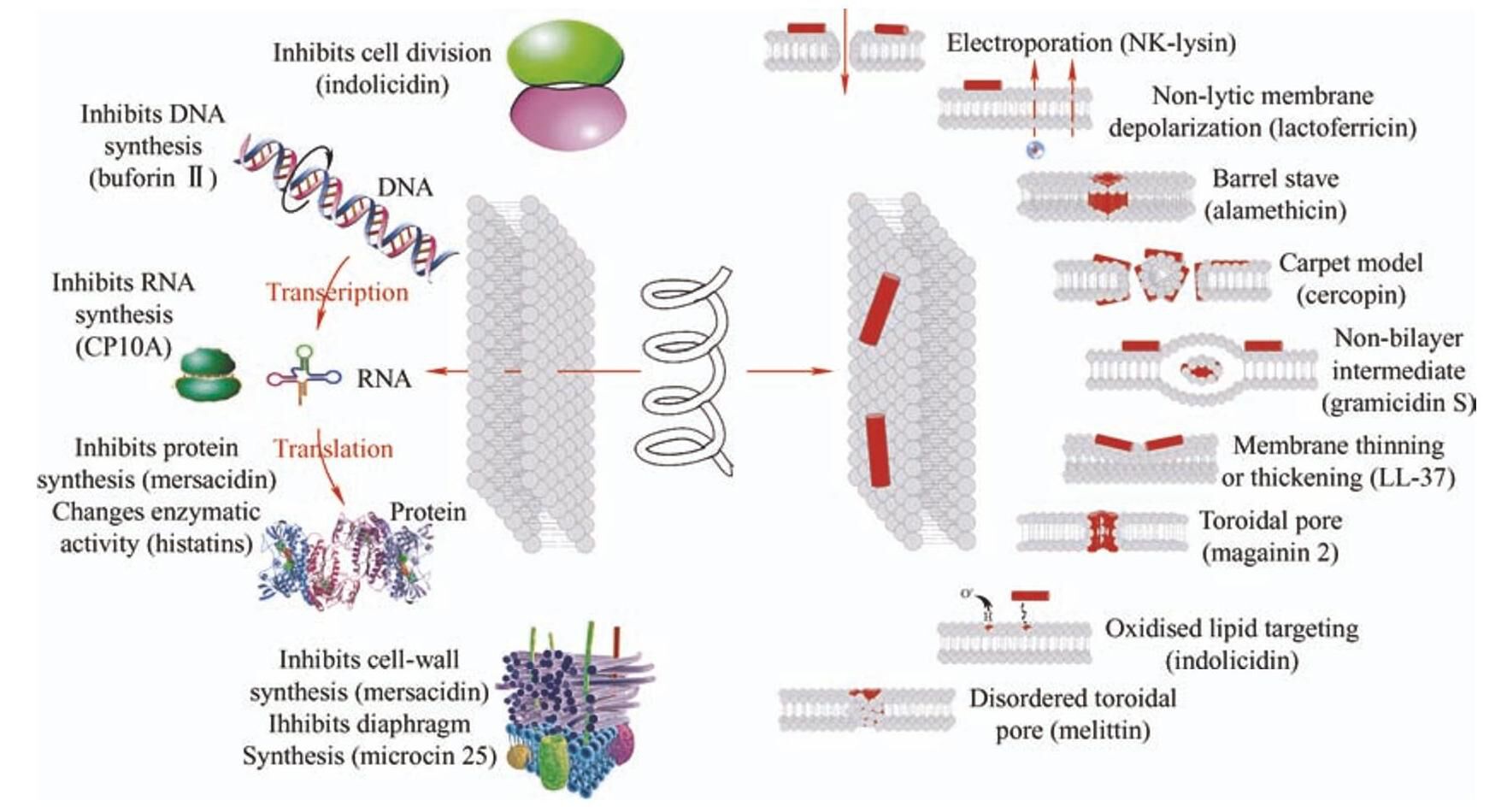
图2 抗菌肽引发的非膜损伤(左)以及膜损伤(右)的多种抗菌机理[16, 18]
2.1 膜损伤
抗菌肽有三种经典的细胞膜损伤的方式:桶板模型、环孔模型和地毯模型。在桶板模型中,抗菌肽聚集并插入膜双层中,疏水性部分向外与脂质核心区域结合,亲水性部分向内构成桶板壁。环孔模型中,抗菌肽同样聚集插入膜中,但极性部分与脂质的极性头基结合,诱导脂质单层膜向内连续弯曲形成环形孔,孔隙由抗菌肽和脂质头基组成,与桶板模型最基本的区别就在于抗菌肽与脂质头基的结合。地毯模型中,抗菌肽被静电吸引覆盖在膜表面的阴离子磷脂头基上,以洗涤剂一样的方式扰乱双分子层,最终导致胶束的形成,同时在临界阈值浓度下,抗菌肽在细胞膜上形成短暂的孔洞,允许额外的抗菌肽进入,以同样的方式破坏另一半分子层,使双分子层因为局部曲率被破坏后解体并形成胶束。
随着对抗菌肽杀菌机理研究的进一步深入,发现了区别于这三种经典模型以外的破坏形式。例如melittin会在膜上形成一种涉及抗菌肽数量更少的无序环形孔[19];短杆菌分泌的一种环十肽gramicidin S可以诱导双分子层形成非层状结构 [20]; Lactoferricin不会破坏膜的完整性,但会诱导膜改变其通透性,允许小离子通过,导致跨膜电化学和pH梯度的损失,造成膜去极化[12];LL-37会改变膜的厚度,但不会在膜上形成孔洞[17];拥有足够高电荷的NK-lysin会在膜上形成电穿孔[21];活性氧会诱导脂质氧化,在细菌膜上产生高亲和力的抗菌肽结合位点[22]。
抗菌肽破坏膜的方式多种多样,学者将单一或多种脂质组合成膜或囊泡来模拟细菌细胞膜,与抗菌肽一起培育,通过各种不同的手段观测抗菌肽与脂质膜相互作用过程中不同的目标对象,以研究特定抗菌肽的抗菌机理[23-24]。研究内容主要包括抗菌肽构象变化、膜孔洞形成、离子通道以及胞内物质改变:
(1)抗菌肽。圆二色光谱和固体核磁共振已经用来测量与脂质膜结合的抗菌肽的取向和二级结构,从而明确抗菌肽破坏膜过程中的活性构象变化[25-26]。
(2)膜孔洞。通过各种高精度显微镜可以直观地观察抗菌肽在脂质膜上形成的孔洞和作用靶点[27]。中子衍射可以记录抗菌肽诱导形成的膜孔的衍射图案,以便准确地测量孔的内外直径[28]。
(3)离子通道。通过监测抗菌肽在膜双层中形成的孔所产生的电流来评估其穿透双分子层的能力以及稳定性[29]。
(4)胞内物质。使用荧光染料对细胞内的球蛋白、右旋糖酐、离子等物质进行染色,再通过荧光显微镜观察胞内物质外泄的情况,可以测定抗菌肽的杀菌时效,研究膜类型以及抗菌肽浓度对抗菌肽杀菌性能的影响[30]。
这一系列手段虽然可以分别监测抗菌肽从结合到破坏膜过程中的某些环节,但无法直观地观察分子水平下细胞膜被破坏的完整动态过程。随着计算机技术的大力发展,分子动力学模拟就像一台超高精度的“摄像机”,以原子分辨率描述系统中所有组件之间的相互作用,能够弥补试验方法的这一部分缺陷。各种不同的仿真算法和分子力场被用来研究肽与膜的相互作用(图3),以期获得更为直观精确的细节[31]。对于肽与膜的模拟主要有两种方式:全原子分子动力学模拟和粗粒度分子动力学模拟。全原子分子动力学模拟是根据牛顿力学来模拟体系的相互作用以及运动情况,然而体系的性质是由大量相互作用的总和决定的,其中一些相互作用很弱,但累积起来有很大的贡献,这远远超出了目前的计算能力,所以需要对牛顿力学进行一些截断以及简化,也就产生了一些不同的力场,如CHARMM、 AMBER、Slipids、GROMOS等,各种力场都有各自的优势,但与实际存在一定的误差。为了增加模拟的时空范围,省略一些影响较小的因素,开发出了低分辨率的粗粒度分子动力学模拟。粗粒度分子动力学模拟通过对结构的简化,根据参考数据的结构特征或者试验数据,演化出Martini、SDK、ELBA、 SIRAH等不同的参数力场,以期在更长的时间和更大的尺度范围内解释肽与膜的相互作用,但其精度远低于全原子分子动力学模拟。虽然目前的分子模拟只可描述一些简单的动态过程,但随着计算机技术的发展以及理论的深入研究,未来有望精准呈现肽破坏膜的全过程。

图3 不同力场下抗菌肽与膜的相互作用模式[31-33]
2.2 非膜损伤
尽管离子通道、跨膜孔的形成和膜破裂最终导致细菌细胞的溶解,然而越来越多的证据表明,膜损伤并不是抗菌肽抗菌的唯一机制。Buforin II等抗菌肽有一些独特的机制可以在不破坏细胞膜的情况下简单有效地转移到细胞质中,当累积到一定浓度后,会作用于核酸、蛋白质、酶等胞内物质,影响细胞的一些关键过程[14-16]。例如:阳离子肽Pep5可以激活细胞壁溶解酶;indolicidin会改变细菌的形态,导致其无法进行细胞分裂;mersacidin可以抑制细胞壁的合成;pleurocidin可以影响DNA、RNA以及蛋白质的合成;histatins可以降低酶的活性; microcin 25可以阻碍细胞隔膜的形成。
抗菌肽的抗菌模式多样,作用靶点从细胞膜到蛋白质、酶、核酸等胞内物质。此外,magainin和indolicidin等抗菌性能优异的肽还采取多种作用模式相互结合,这增加了抗菌机理研究难度的同时,也使得细菌不易对抗菌肽产生如对抗生素一样的耐药性。同种抗菌肽多种抗菌模式结合的复杂机制意味着高抗菌效率、稳定的抗菌性能以及易规避细菌的耐药性,使抗菌肽的长久应用成为可能[34-35]。
3 抗菌肽在基体表面的接枝方法
要将抗菌肽应用于海洋防污领域,必须实现稳定高效的基体表面接枝方法。
3.1 自发接枝
自然界中有些抗菌肽可以自发的与表面结合,最常见的是肽中半胱氨酸残基(Cys)形成的二硫键与表面共价结合[36]。XIAO等[37]设计了一种天蚕素蜂肽混合的抗菌肽,通过羧基端的Cys,将其固定到两种不同的有机涂层表面:① 通过化学气相沉积(CVD)制备的二溴马来酰亚胺聚合物表面; ② 通过叠氮-聚乙二醇-马来酰亚胺分子与炔基膜反应制备的马来酰亚胺自组装单层膜表面。研究发现该肽通过CVD法制备的聚合物表面的稳定性能、抗菌性能优于自组装单层膜表面。除了含有Cys的肽外,许多肽对金属、氧化物、有机物等材料表面都显示出很强的亲和力,这些肽被认为可以识别材料表面原子或分子的三维结构[38]。然而,同时具备抗菌功能片段和表面亲和片段的天然肽很少。使用戊二醛作为交联剂或者通过卤素原子的亲核/亲电取代反应可以在天然抗菌肽中引入二硫键,从而使其拥有表面亲和能力[39]。通过基因工程技术来获得期望功能的改性抗菌肽是目前普遍采用的方法,可以很好地弥补这一部分抗菌肽的空缺。NILEBACK等[40] 通过基因融合的方法将magainin I重组结合到丝蛋白上,这种丝蛋白可以在各种表面自组装成纳米纤维结构,而magainin I片段可以起到杀菌作用,所形成的涂层可以在氯化氢、氢氧化钠和乙醇溶液的洗涤下保持稳定。
3.2 偶联剂接枝法
除了通过抗菌肽自身特殊片段的结合能力之外,还可以利用一些偶联剂将肽间接固定到基体表面上。常用的偶联剂有多巴胺、聚乙二醇和糖类聚合物等。
3.2.1 多巴胺
多巴胺是一种功能性极强的偶联剂,其反应性儿茶酚基团可以通过亲核Michael加成或Schiff碱反应结合抗菌肽。多巴胺在有氧、碱性条件下进行氧化聚合自组装成粘附力极强的聚多巴胺薄膜[41]。 LOU等[42]通过多巴胺将nisin结合到具有微结构的玻璃表面,这种物理生物协同的防污表面显示出优异的防污性能。MAITY等[43]设计了一种可以在金属表面自组装并拥有防污功能的三肽结构,该肽通过3, 4-二羟基-苯丙氨酸(DOPA)固定到表面上,通过其余两个苯丙氨酸残基的 π-π 堆叠引导肽在表面自组装形成有序的超分子结构。同时,对苯丙氨酸残基的芳香环进行氟化改性,形成一种疏水氟化聚合物。DOLID等[44]设计了一种拥有自组装、抗菌等功能的六肽,通过其氨基末端的DOPA在聚二甲基硅氧烷(PDMS)表面自组装形成稳定的防污涂层。然而,这只是一种简便的肽结合策略,无法准确地调控肽的各种性能。
3.2.2 聚乙二醇
聚乙二醇(PEG)是一种亲水性聚醚,被认为是最好的生物相容性聚合物之一[45]。PEG可以通过与肽结构中的酰胺、硫醚和三唑等基团发生反应,将肽共价固定到基体表面[46-47]。NIE等[48]通过PEG与KR-12上的酰胺基团发生反应,将KR-12共价结合到钛表面。与KR-12直接结合到表面相比,通过PEG连接的KR-12抗菌效率更高。CHEN等[49]通过PEG与硫醚的连接,将daptomycin共价结合到钛合金Ti6Al4V表面。与简单的多巴胺结合策略相比,采用PEG连接有三个很明显的优点:①使表面具有亲水性,增加防污能力;②允许抗菌肽的横向扩散和构象转变,保证抗菌活性;③控制抗菌肽的释放,提高稳定性能。
3.2.3 糖类聚合物
透明质酸、丹宁酸、壳聚糖等糖类聚合物也可以作为偶联剂将抗菌肽结合到基体表面。YE等[50] 首先在金表面化学接枝上硫代透明质酸,然后通过两性离子多肽p-EK的自组装功能得到多肽改性表面。该表面具有很强的亲水性,同时抗蛋白性能优于只用透明质酸修饰的表面。XU等[51]利用肽parasin I和丹宁酸之间的Michael加成/Schiff碱反应,将这两种材料交替沉积在海军应用的不锈钢表面上,制备了多层防污涂层。随着膜层数量的增加,膜层的防污性能和抗菌性能均有所提高。天然海水浸泡15d后,涂层仍表现出很好的稳定性和抗菌性。这种层层自组装的策略优点在于能便捷地控制涂层中肽的含量,难点在于涂层的稳定性与通透性之间的平衡。一方面,涂层包裹的肽想要暴露在环境中起到抗菌功能,需要透过连接层,或者连接层能够自发的水解消耗掉。另一方面,肽释放的速度或者连接层水解的速度不能过快,以确保防污效果的长久性。解决了这两方面矛盾以后,层层自组装的涂层才可以具有长久稳定的防污能力。
3.3 偶联剂-缓冲片段接枝法
抗菌肽的抗菌活性依赖于合适的空间结构,因此在接枝过程中需要尽可能地保持其二维结构。在偶联剂与抗菌肽之间嵌入一个缓冲片段,可以维持与抗菌活性相关的肽结构基序,允许抗菌功能片段的外露和构象变化的灵活性,更接近溶液中抗菌肽的行为特性[52]。最常见是在肽链中加入一个额外的半胱氨酸,利用半胱氨酸上的巯基与偶联剂上的硫醇、马来酰亚胺等基团的反应,将肽共价结合到基体表面。COSTA等[53]在Ti和Au表面沉积壳聚糖薄膜,随后用N-乙酰半胱氨酸在薄膜上引入巯基。 Dhavar5通过其上的半胱氨酸与预功能化膜上的巯基之间形成二硫键,共价结合到表面上。YAZICI等[54]将一个由三个甘氨酸残基组成的柔性片段嵌入表面结合肽与抗菌肽之间,结果表明抗菌肽通过柔性片段固定到表面后,保持了其三维螺旋结构。同时有研究表明,与柔性肽连接剂(GGGGS)相比,刚性肽连接剂(PAPAP)显著增加了嵌合肽在钛表面的吸附[55]。
抗菌肽在基体表面的固定是应用于防污领域的前提。肽的固定能够通过相对随机的方式进行,例如,通过肽的胺基和基体表面上的羧基形成酰胺键。另外,也可以通过化学选择性形成特定的共价键,使其以一种可预测和确定的方式结合到基体表面。与随机结合相比,固定控制策略更可取,因为抗菌肽的抗菌活性取决于其特定的氨基酸片段和空间结构。随机固定的方式可能会破坏抗菌肽的活性构象或者片段,使抗菌性能降低。最理想的固定控制策略是抗菌肽在基体表面体现出其在溶液中的特性,这需要根据抗菌肽的序列、空间构象以及活性片段对化学耦合策略进行多方面的调控。
4 抗菌肽的海洋防污性能
抗菌肽对抗海洋污损的手段多样,除了前文提到的抗菌功能以外,还能影响生物膜的形成,例如, Nisin可以降解芽孢杆菌形成的生物膜[56],海洋雪蟹分泌物中提取出来的抗菌肽可以减少低碳钢表面的生物膜[6]。抗菌肽主要从以下两个方面影响生物膜的形成。
(1)降低胞外聚合物(EPS)[56]。生物膜中胞外聚合物的含量高达90%以上,胞外聚合物由多糖、核酸、蛋白质以及脂类等物质组成。Nisin可以降低胞外聚合物中的多糖和eDNA含量[57]。Hepcidin-20可以作用于多糖胞间黏附素,降低胞外基质含量,改变生物膜的构架组成[58]。S4(1-16)M4Ka通过分解脂质、分散细菌来抑制生物膜的形成[59]。Piscidin-3可通过N端氨基酸与Cu2+形成共价键,发挥核酸酶活性,破坏胞外DNA[60]。
(2)调控信号分子[61]。细菌会对周围的环境做出严格的应答性反应,释放信号传递分子[62]。鸟苷四磷酸和五磷酸是一种普遍存在各种细菌中的信号分子,可以调控大量与蛋白质合成相关的基因表达[63],能够控制芽孢杆菌、假单胞菌、链霉菌以及弧菌等属细菌的生物膜形成过程[64-66]。抗菌肽可以影响鸟苷四磷酸和五磷酸的产生,阻断细菌的应答性反应信号的传递,从而阻止生物膜的形成[67]。群体感应信号分子是另一类调控细菌行为模式的物质[68]。群体感应信号分子在环境中不断积累,当浓度达到阈值以后,调节蛋白就会被触发并进一步诱导目标DNA,导致群体感应调节基因的转录,进而改变细菌的社会行为。这类信号分子密切调控着生物膜的形成[69]。LL-37能够通过抑制关键信号分子的传递,阻止50多个与生物膜形成相关的基因表达[70]。
抗菌肽同样可以抑制微藻的附着。BEYER等[71] 制备了一系列抗菌肽自组装单层膜,可以有效减少舟形藻的附着,防污效果优于疏水性的正十二硫醇表面和亲水性的聚乙二醇表面。此外,他们在金表面制备的其他系列抗菌肽自组装薄膜,同样可以减少77%~97%的舟形藻附着[72]。CAO等[73]在具有微结构的304不锈钢表面接枝抗菌肽,对小球藻和三角褐指藻的附着抑制率分别达到78.56%和87.80%。乳铁蛋白类的抗菌肽会影响新月菱形藻、双眉藻等微藻的生长和附着[74],但抗菌肽减少藻类附着的具体作用机制还不够明确。
污损生物群落中除了细菌和微藻这些小型污损生物以外,大型污损生物是群落中最棘手的成员,也是船体和海上平台的主要破坏者。大型污损生物附着在基体表面前会经历一个自由生活的浮游扩散阶段,称为繁殖体(幼虫和孢子等)。对于大多数大型污损生物来说,繁殖体所选择的基体对随后的生存和繁殖特别重要,一旦幼虫或孢子附着在基体上,它们就会蜕变为幼体并收集能量,迅速或缓慢地成长为成体,终生不会改变位置。肽在海洋系统中具有很高的溶解度,半衰期较短,具有较高的信噪比,是很好的信号分子。肽的氨基酸序列就和文字排列一样,不同的序列传达出不同的信号,可以作为一种信息素来影响大型污损生物的附着[75]。
藤壶是一种常见的海洋污损生物,群体附着在船体和其他浸入水中的物体表面,也是目前研究最多的大型污损生物模型。STEFFEN等[76]发现从深海海绵中提取的抗菌肽barrettide C可以明显地抑制藤壶幼虫的附着,但只会破坏幼虫的运动技能,而不会杀死幼虫。EDERTH等[77]为了调查抗菌肽对藤壶幼虫附着的影响,设计了四种基于赖氨酸和谷氨酸交替沉积的两性离子抗菌肽自组装膜。结果表明,只有一种以谷氨酸终止,不包含碱性氨基酸,而且不能被丝氨酸蛋白酶降解的抗菌肽能有效阻止藤壶幼虫的附着。同时发现,以赖氨酸或精氨酸结尾的电中性、碱性三肽是比较典型的信号肽结构[78]。这类三肽的一个例子是甘氨酸-甘氨酸-精氨酸(GGR),它是甲壳类和软体动物的一种肽类信息素[78]。这种信息素是通过胰蛋白酶类的丝氨酸蛋白酶对有机基质中的蛋白或者天然胶降解产生,可以影响藤壶幼虫的附着。贻贝同样是普遍存在的大型污损生物,是污损生物群体中的重要组成部分,海洋海绵动物Geodia barretti中提取的环肽对贻贝幼虫的附着和变形有强烈影响[79]。 SJOGREN等[80]从蜂海绵中提取出一种能够驱赶贻贝幼虫的六肽。ANTUNES等[81]从海洋蓝细菌中提取出环肽portoamides可以有效阻止贻贝幼虫的附着,但在任何浓度下都不会杀死贻贝幼虫,这表明该肽通过信号传递功能赶走贻贝幼虫。肽同样会影响石莼孢子的附着。SILVA-ACIARES等[82]在智利北部发现一种细菌的胞外产物中存在一种抗菌肽,可以抑制石莼孢子的附着。CALABRESE等[83]合成的一种寡肽同样可以有效地防止石莼孢子的附着。EDERTH等[84]发现富含精氨酸的抗菌肽会改变石莼孢子的附着方式,不经过常规的表面勘探,分泌粘合剂等过程,直接附着到基体表面。这些肽对石莼孢子的影响是通过特定的肽-膜相互作用介导的。抗菌肽作为信号素来防止大型生物繁殖体附着的功能主要体现在序列上。抗菌肽序列直接决定了其功能:抑制附着、诱导附着或是改变附着模式等。读懂序列中的隐藏信息,建立序列和信号的关系模型是抗菌肽在防止大型生物污损领域成功应用的前提。
5 结论与展望
自20世纪80年代抗菌肽发现以来,因其广谱抗菌活性、低浓度快速杀菌能力和环境友好性而成为防污领域的研究热点。本文对抗菌肽在海洋防污领域的应用进行了全面总结,主要有以下结论。
(1)抗菌肽的杀菌活性依赖于其稳定的二级结构,主要包括 α-螺旋结构、β-折叠结构和延伸/弹性结构。
(2)抗菌肽拥有多种杀菌模式,一方面通过多种方式破坏细菌细胞膜,另一方面作用于酶、蛋白质、核酸等胞内物质,影响细菌细胞的一些关键过程。
(3)抗菌肽除了通过自身特殊片段的结合能力之外,还可以利用多巴胺、聚乙二醇、糖类聚合物等偶联剂间接固定到基体表面上。然而,不同的接枝方法对抗菌肽的性能影响很大,在偶联剂与抗菌肽间加入缓冲片段有利于抗菌肽保持其抗菌特性。
(4)抗菌肽不仅拥有直接杀菌的特性,还能够减少环境中污损生物的浓度、抑制生物膜的形成、影响生物间信号分子的传递,通过多种方式共同作用有效地抑制生物污损。
虽然目前抗菌肽在海洋防污领域的应用研究取得了很多标志性成果,但还存在一些须要攻克的难题,也决定了未来抗菌肽的发展方向,主要体现在以下几个方面:
(1)抗菌肽制备的未来发展方向。目前获得抗菌肽的方法主要有五种:①直接从天然生物体内提取抗菌肽,但由于生物体内抗菌肽含量很低,存在提取效率低、成本高的问题;②根据已知的抗菌肽序列,化学合成抗菌肽,但只适用于序列较短的肽; ③通过酶水解蛋白获得抗菌肽,但水解副产物的存在,使得目标抗菌肽的分离难度大,浓度低;④通过基因工程的方法获得抗菌肽,但技术要求高,成本相对昂贵;⑤利用细菌产生细菌素或肽类毒素获得抗菌肽,但同样存在技术难度大,造价昂贵的缺点。如何大量、低成本、高效率的制备目标抗菌肽是未来的一个重要发展方向。
(2)抗菌肽接枝方法的未来发展方向。虽然自然界中有些抗菌肽可以自发的与基体表面结合,但同时具备抗菌功能片段和基体亲和片段的天然肽很少。一般都通过偶联剂将肽间接固定到基体表面。但不同的化学耦合策略对抗菌肽的构象和活性影响很大。如何确保抗菌肽固定到表面后还能体现出和在溶液中一样的空间结构、抗菌特性是另一个发展方向。
(3)抗菌肽的构效关系研究是实现抗菌肽广泛应用的重要前提。抗菌肽的性能依赖于一个稳定的活性构象。同时,不同序列抗菌肽所表现出的活性构象、抗菌机理以及防污性能大不相同。明确序列、结构与功能之间所存在的关系是抗菌肽广泛应用的重要前提。
参考文献: 略
免责声明:本网站所转载的文字、图片与视频资料版权归原创作者所有,如果涉及侵权,请第一时间联系本网删除。
相关文章

官方微信
《腐蚀与防护网电子期刊》征订启事
- 投稿联系:编辑部
- 电话:010-62316606-806
- 邮箱:fsfhzy666@163.com
- 腐蚀与防护网官方QQ群:140808414
点击排行
PPT新闻

“海洋金属”——钛合金在舰船的
点击数:8156

腐蚀与“海上丝绸之路”
点击数:6487