
微生物腐蚀机理研究进展
2016-07-27 11:47:07
作者:张盾 吴佳佳 王鹏 中国科学院海洋研究 来源:《腐蚀防护之友》
分享至:


微生物腐蚀简介
微生物腐蚀的定义
微生物腐蚀是指有微生物参与的腐蚀,它有几个显著特点:首先,微生物腐蚀是一电化学过程;其次,微生物能够影响腐蚀的程度和过程;再次,微生物腐蚀的发生除需要微生物外,还需要水、能量来源、碳源、电子供体、电子受体等。微生物腐蚀几乎可在所有环境中发生,由其造成的损失约占腐蚀总损失的 20%。
微生物腐蚀的研究发展历程
参照人类社会发展史,微生物腐蚀的研究历史也可划分为古代、近代和现代三个时期。19 世纪中叶到 20 世纪 20年代可称为微生物腐蚀研究的“古代”,在此期间人们意识到了微生物参与并影响腐蚀过程,而且确定了硫酸盐还原菌(SRB) 在微生物腐蚀中的重要性。在 20世纪 20 年代到 60 年代的“近代”,出现许多里程碑式的研究结果,如氧化还原电位可作为微生物腐蚀的重要指示、氢化酶的发现、首例由微生物腐蚀导致的地下管道破坏事故的确定、SRB 经典阴极去极化理论的提出等。20 世纪 80年代以后,随着不同学科之间交流的加强以及研究手段的不断进步与融合,微生物腐蚀研究报道在数量和质量上有了突跃发展,人们对微生物腐蚀机理的认知也在不断深化,由此开启了“现代”时期。
微生物腐蚀所涉及微生物的复杂性
在自然环境中,腐蚀金属表面往往覆盖有生物膜,生物膜可以为微生物的生存生长繁殖提供保护。在生物膜的不同深度,溶解氧、有机物、无机盐等的浓度不同,使得分布在不同阶层的微生物种类存在差异 ( 图 1)。在接近流体相的生物膜表层,溶解氧含量高、营养物质供给方便,该层的微生物进行有氧代谢,降解有机物产生简单聚合物和有机酸,硝化细菌、锰氧化菌、铁氧化菌(IOB)、硫氧化菌 (SOB) 等均可在此层出现。随着生物膜深度的增加,溶解氧浓度降低,可以利用来自表层微生物的代谢产物的兼性厌氧菌得以生存,如某些进行发酵、反硝化代谢的微生物。在靠近金属基体的生物膜底层,溶解氧浓度非常低、处于贫氧状态,这利于厌氧微生物的生存,包括甲烷菌、SRB、铁还原菌 (IRB) 等。
在以上提及的与腐蚀有关的微生物中,SRB、SOB、IOB、IRB 对腐蚀的影响受到人们的重视,随着研究工作的深化,其腐蚀机理也不断得到发展。
典型腐蚀微生物的腐蚀机理
SRB
SRB 长期以来被认为是最重要的厌氧腐蚀微生物,由其造成的腐蚀损失约占整个微生物腐蚀损失的一半以上。
SRB 的突出特征为可将硫酸盐还原为硫化物从中获取能量。自 1934 年经典去极化理论的提出,目前被报道的 SRB 腐蚀机理有多种,下边分别给以介绍。
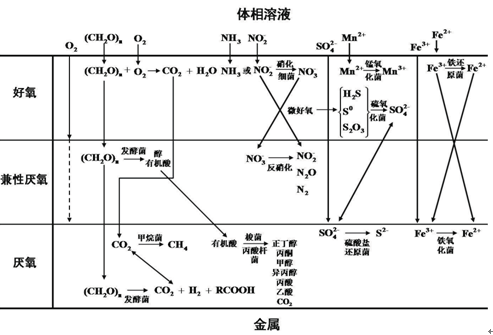
图1 腐蚀金属表面典型生物膜的层状特征及可能的反应
(1) 经典去极化理论
该理论的核心思想为 SRB 通过氢化酶的作用移除金属表面的吸附氢用于硫酸根还原,从而加速腐蚀。在阳极区和阴极区分别发生铁的溶解 ( 反应式 (1)) 和析氢反应 ( 反应式 (2)),阴极反应形成的吸附氢阻碍了反应的进一步进行;而 SRB 在氢化酶的作用下可将吸附氢利用 ( 反应式 (3)),实现阴极的去极化,使得反应得以继续进行;阴极去极化产生的 S 2- 可与阳极溶解产生的 Fe 2+ 结合形成 FeS 腐蚀产物 ( 反应式 (4)),由于 SO4 2- 与 Fe 的反应比为 1 : 4,那么多余的 Fe 2+ 与水电离产生的 OH - 结合形成 Fe(OH) 2 ( 反应式 (5));总的反应式如式 (6) 所示。

虽然经典去极化理论第一次从电化学的角度解释了 SRB 对腐蚀的影响,但该理论的合理性和适用性受到了人们的质疑。例如:有的研究指出氢化酶不能对原子氢起作用;根据该理论,腐蚀的 Fe 与生成的 FeS 的摩尔比为 4 : 1,但在实际情况下,比值为 0.9-1;有的 SRB 可以还原硝酸盐,当硝酸盐存在时,SRB 能够有效氧化金属表面的阴极氢,但并未导致腐蚀加速。
(2) 硫化物的去极化作用
该理论认为 SRB 作用下产生的金属硫化物的氧化还原电位高于金属基体的,这样就会形成以金属基体为阳极、金属硫化物为阴极的腐蚀原电池,从而导致腐蚀速率的增加。
(3) 挥发性磷化物的加速作用
该理论认为 SRB 在生长代谢过程中可以产生腐蚀性强的磷化物 ( 如 H 3 P),其可与 Fe 反应,从而加速腐蚀。
(4) 阳极去极化理论
该理论认为 SRB 利用阴极反应产生的 H 2 将 SO4 2- 还原产生 H 2 S,H 2 S 电离产生 S 2- 和 H + 离子,而 S 2- 消耗阳极溶解产生的 Fe 2+ 生成 FeS,进而加速阳极的活化溶解。该理论与 (1) 中所示经典去极化理论的主要差别在于,前者认为Fe 的阳极溶解是速率控制步骤,而后者认为吸附氢的移除是速率控制步骤。
(5) 胞外多聚物螯合金属离子理论
SRB 在金属表面的附着生长繁殖形成生物膜,胞外多聚物是生物膜的重要组成部分。该理论认为某些 SRB 产生的胞外多聚物能够与金属离子螯合,从而加速金属的阳极溶解。
(6) 应力腐蚀开裂理论
该理论认为 SRB 产生的 H 2 S 能够降低局部微环境的 pH、抑制沉积型腐蚀产物的形成,从而促进氢的渗透、降低金属材料的弹性,导致腐蚀应力裂纹的生长和氢脆。
(7) 硫化物的生物矿化理
论该理论认为 SRB 生物矿化作用产生的硫化物使得某些金属材料的腐蚀电位负移,从而使得腐蚀速率增加。
(8) 三阶段理论
该理论将 SRB 作用下的腐蚀划分为三个阶段。第一个阶段受菌体细胞的吸附和硫化物 ( 主要是马基诺矿和黄铁矿 ) 在材料表面的沉积控制,由硫化物作为阴极、基体金属作为阳极构成的微腐蚀原电池使得氢渗透峰出现。在第二个阶段,细菌和无机物处于平衡状态,由硫化物和胞外多聚物组成的较为致密的膜层使得金属的开路电位略有正移。在第三个阶段,SRB 的代谢活性使得局部 pH 降低,黄铁矿向马基诺矿转变,导致严重的局部腐蚀,并发生腐蚀产物和生物膜的脱落。在第三个阶段,腐蚀严重,但由于胞外多聚物的阻碍作用没有氢渗透的发生。
(9) 直接获取电子理论
该理论针对特定的自养型 SRB 物种,它们可以导致腐蚀速率的显著增加,而不需要消耗氢气。SRB - 方面通过与金属材料的直接接触获取来自阳极 Fe溶解释放的电子,另一方面伴随着 FeS腐蚀产物的积累,SRB 与基体金属材料之间被 FeS 隔离,但由于 FeS 是良好的电子导体,因而附着在和包裹于 FeS 的SRB 依然可以直接获取来自金属基体氧化释放的电子 ( 图 2)。与其它 SRB 物种作用下的腐蚀产物相比,该理论下的腐蚀产物层主要以无机硬质形式存在,而不存在大量的菌体细胞和胞外多聚物。
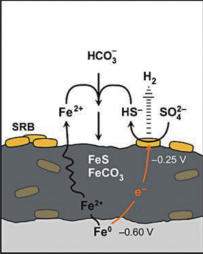
图2 SRB直接获取电子理论示意图
(10) 生物催化阴极硫酸盐还原机理
该理论从生物能量学角度出发,认为 SRB 加速腐蚀的原因为其需要从金属基体上获取电子来还原 SO4 2- 获取能量维持生存 ( 图 3)。与其它传统理论不同,该理论没有固定的物理阴极,而是将 SRB 作为生物阴极。
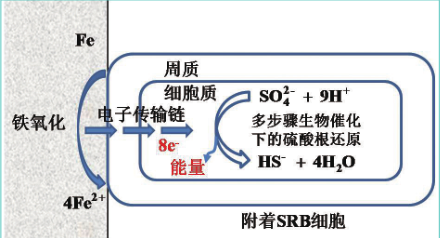
图3 生物催化阴极硫酸盐还原机理示意图
需要注意的是,该理论只适用于那些与金属基体直接接触的菌体细胞,而非整个 SRB 生物膜。
以上腐蚀机理的提出,均针对厌氧体系。纵观 SRB 腐蚀机理的发展史,可以看出其经历了由突出微生物的作用( 理论 (1)) 到弱化微生物的作用 ( 理论(2)-(7)),再到重新认识到微生物的重要作用 ( 理论 (8)-(10)) 的过程。这些腐蚀机理均有一定的合理性与适用范围,根据环境条件的不同,一种或多种机理可用于解释特定体系 SRB 对腐蚀的影响。
(11) 有氧条件下 SRB 对腐蚀的影响
在实际环境中,SRB 往往可从有氧条件下形成的生物膜中分离得到,生物膜中其它好氧微生物的代谢可为 SRB 的生长提供厌氧条件,同时,SRB 也可能与氧直接接触。那么,有氧条件下 SRB是否对腐蚀有影响?如果有影响的话,影响机制是怎样?
人们在一些 SRB 物种中发现了耐氧蛋白,然而 SRB 能否利用氧作为电子受体依然存在争论。某些 SRB 能够表达过氧化物酶、超氧岐化酶等,而这些酶可能对金属表面的溶解氧阴极还原反应有影响,进而影响腐蚀过程。因而,我们研究组前期围绕有氧条件下 SRB 对碳钢腐蚀的影响及 SRB 对溶解氧还原反应的影响做了大量工作。已有研究结果表明相比厌氧环境,氧的引入使得 SRB 生长受到抑制、介质中 S 2- 的浓度较低,生物膜结构疏松且出现局部团聚现象,在与溶解氧的共同作用下,使得碳钢腐蚀加速;SRB 的加入能够抑制碳钢上溶解氧还原反应的进行,且不同的代谢产物对溶解氧还原反应的影响不同,如硫化物、葡萄糖醛酸抑制该电化学反应,而D- 甘露糖则起促进作用。这些研究结果证实了有氧条件下 SRB 所致腐蚀与厌氧条件下的存在差异,同时,SRB 对溶解氧还原反应的影响可能影响有氧条件下的腐蚀。
SOB
SOB 的突出特征为利用硫化物、单质硫、硫代硫酸盐、亚硫酸盐等还原态硫化物氧化为硫酸的转变获取能量。
一般认为,SOB 通过酸腐蚀机理加速材料的腐蚀,其代谢产生的硫酸能够降低 pH、不利于某些保护性腐蚀产物的沉积,从而促进腐蚀的发生。目前,对SOB 的腐蚀研究主要集中于金属材料的加速低水位腐蚀和混凝土的加速腐蚀。
当 SOB 与 SRB 共同存在时,SOB 产生的SO4 2- 可被 SRB 利用,而 SRB 产生的 S 2-又被 SOB 利用,如此实现硫循环,可导致更加严重的腐蚀。
IOB
IOB 利用 Fe(II) 到 Fe(III) 的氧化获取能量,其往往被认为与管道中锈瘤的形成密切相关。
IOB 生物膜及 Fe(III) 腐蚀产物在金属表面的沉积使得被覆盖部分溶解氧浓度降低成为贫氧区,而未覆盖部分溶解氧含量高作为阴极区,小阳极大阴极的结构使得覆盖区电流密度大。阳极溶解产生的金属离子发生水解产生 H + 使得局部 pH 降低,同时,大量 H + 的积累引起体相溶液中腐蚀性阴离子 ( 如 Cl - ) 向阳极区迁移,从而诱发点蚀。同时,IOB生物膜和 Fe(III) 腐蚀产物沉积所形成的锈瘤为 SRB 等厌氧微生物的生长提供了便利,多种微生物的共同作用使得腐蚀速率进一步增大。
IRB
IRB 是 一 类 具 备 将 Fe(III) 还 原 为Fe(II) 能力的微生物,从对氧的需求角度来讲,IRB 包括严格厌氧菌和兼性厌氧菌,在流动体系氧被频繁引入的情况下,兼性厌氧菌比严格厌氧菌更有竞争优势。
IRB 对腐蚀的影响既可以是促进,又可以是抑制,这与菌株、测试体系性质等密切相关。图 4 给出了一兼性厌氧IRB 对腐蚀过程影响的示意图。在腐蚀初期,溶液介质中的溶解氧含量高,IRB在金属材料表面的不均匀附着导致局部溶解氧浓度降低,溶解氧的浓度梯度导致局部阴阳极的出现,引发腐蚀加速(A)。随着游离和附着 IRB 生长对氧的消耗,金属表面溶解氧浓度梯度消失,因而局部阴阳极消失;一旦金属表面氧被耗尽,兼性厌氧 IRB 转向以 Fe(III) 为电子受体,产生的 Fe(II) 向体相溶液扩散;Fe(II) 容易与溶解氧反应使得溶解氧浓度进一步降低,腐蚀被抑制 (B)。IRB 的腐蚀抑制作用在静态体系中比较明显,因为 Fe(II) 得以大量积累,溶解氧难以到达金属表面 (C)。在流动体系中,一方面,产生的 Fe(II) 被连续带走,其浓度较低;另一方面,氧被持续输入,再次出现局部阴阳极,腐蚀被促进 (D)。
除由氧浓差电池造成的腐蚀促进外,IRB 对腐蚀的促进作用还可以通过将金属表面具有保护性的 Fe(III) 氧化层破坏来实现。
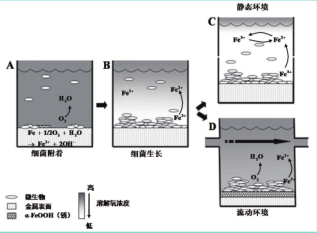
图4 兼性厌氧IRB对腐蚀过程的影响
结束语
尽管我们已对微生物在腐蚀中的作用进行了大量研究,但我们必须谦逊地承认这些微小的生物体的确具备使我们迷惑的能力。以上所提及的腐蚀机理在特定的体系有一定的合理性,而在实际环境中,多种微生物并存,他们之间的相互影响使得微生物腐蚀非常复杂。与对微生物腐蚀的未知相比,我们现阶段的已有认知只是沧海一粟。因此,为了推进对微生物腐蚀更深层次的认知,必须投入更大的人力物力财力。
作者简介

张盾,理学博士,中国科学院海洋研究所研究员、博士生导师。中国腐蚀与防护学会副理事长和中国海洋湖沼学会理事、中国海洋湖沼学会海洋腐蚀与污损专业委员会主任。2006 年中国科学院“百人计划”入选者,中国科学院海洋环境腐蚀与生物污损重点实验室主任。现从事海洋环境腐蚀与生物污损的研究,围绕海水的微生物腐蚀与控制问题开展了海水中典型腐蚀微生物与腐蚀过程电化学反应作用机理的研究,研发了一系列基于腐蚀微生物的抗体、代谢过程和遗传片段的生物传感器,开发了具有防腐抗污性能的仿生超疏水和超滑材料、绿色光催化杀菌剂和分子容器防污缓蚀剂。发表论文 200 余篇,申请及授权国家发明专利 50 余项。
更多关于材料方面、材料腐蚀控制、材料科普等方面的国内外最新动态,我们网站会不断更新。希望大家一直关注中国腐蚀与防护网http://www.ecorr.org
责任编辑:邢云辉
免责声明:本网站所转载的文字、图片与视频资料版权归原创作者所有,如果涉及侵权,请第一时间联系本网删除。
相关文章

官方微信
《腐蚀与防护网电子期刊》征订启事
- 投稿联系:编辑部
- 电话:010-62316606-806
- 邮箱:fsfhzy666@163.com
- 腐蚀与防护网官方QQ群:140808414
点击排行
PPT新闻

“海洋金属”——钛合金在舰船的
点击数:8135

腐蚀与“海上丝绸之路”
点击数:6475